Lack of Objective and Robust Methodology
The author presents no systematic, objective and rigorous methodology for determining what "kinds" are. Friar does present a list of guidelines for determining what holobaramins are (hereafter "kinds" and holobaramin will be used interchangeably) but this list is tantamount to a list of features that might be important – similarities in form, ontology, ecology, fossils and the ability to hybridize.
But Friar provides no systematic methodology for how one determines holobaramins, and offers no explanation of how or why each of the data sources are relevant or important. Friar seems to imply that it is up to the jurisprudence of the creationist conducting the study:In order to determine baraminic distances among types of organisms it is important to utilize the most significant data.
There is no mention of how one decides which data are most significant, no objective or quantitative criteria for judging the significance of available biological and ecological data on the organisms in question, and no mention of whether methods exist for assessing the reliability of the results once analysis has been conducted (such as bootstrap or jackknife replicates). Instead, Friar lists several "studies" published in the Creation Science Research Quarterly Journal where the authors assess the relative importance of some data over others, apparently after the study was conducted. Friar concludes:
In particular, proteins, and their DNA patterns, which are part of fundamental cellular processes, have wide applicability in baraminological research. For example, proteins associated with cellular respiration like cytochrome C and cytochrome B are excellent candidates.
One has to wonder what "DNA patterns" are – presumably DNA sequences associated with proteins. Nevertheless, Friar does not cite any study supporting this claim, nor does he indicate how or why these proteins would be important. In short, this paper provides no useful information on how one determines what organisms are members of "kinds". He provides a shopping list of characters that might be useful, but provides no methods for applying them objectively, no indication that the methods are described and tested elsewhere (except anecdotally) and no way to assess the reliability of the results of their analysis.
In contrast, the methods, phylogenetic methods and models, are thoroughly and robustly tested and their efficacy judged by the scientific community. There are textbooks written on the methods of phylogenetic analysis (e.g. Hillis et al. 1996). Software is written, tweaked and tested (e.g. Excoffier et al. 2005; Rambaut & Grassly 1997). Assumptions of methods and models are described, tested and reviewed in the scientific literature (e.g. Weiss & von Haesler 2003) and papers published invoking phylogenetic methodologies must describe the analyses they conducted in considerable detail (see virtually any published article on molecular phylogenetics; e.g. Saux et al. 2004). In fact, papers submitted for publication in scientific journals are routinely rejected because their analyses are inadequate or incomplete. Even though Friar's paper is likely intended as a review paper and not a research paper per se, it is still lacking in any detail on methods and assumptions upon which the efficacy of the model can be judged.
Failure to Qualify as a Scientific Hypothesis
Friar states that:
The goal of baraminology is to characterize holobaramins.
The goal then, is to identify Biblical "kinds" and to describe all of the organisms comprising them. So creationists begin with the assumption that "kinds" exist, in light of scientific evidence to the contrary. They have not tested – nor do they ever intend to test – the question of whether holobaramins actually exist. In fact, Friar deliberately avoids listing the assumptions of their model entirely. The reason is obvious when you read the "guidelines" for practicing bariminology:
Scripture claims (used in baraminology but not in discontinuity systematics). This has priority over all other considerations.
So scripture tells them that "kinds" exist, and they have only to describe them. This is a critical piece of information – it means that they will never, ever admit that "kinds" do not exist. Ever. This is contrary to the way science is conducted – a scientific theory or hypothesis must be potentially falsifiable. That is, there must be a test or tests that, if confirmed, would deny the validity of the hypothesis or theory, and scientists must be willing to part with hypotheses if enough evidence exists that rejects them. In phylogenetic analysis, tree building methods are judged based on, among other criteria, how well they perform when assumptions are violated and whether they alert us when assumptions are violated (Penny et al. 1992) But creationists have no intention of ever testing the vital assumption that "kinds" exist, and since scripture has priority over all other considerations, it is evident that they would dismiss any evidences suggesting that "kinds" do not exist.
Analyses Indicate That "Kinds" Do Not Exist
Is there a test that could deny the existence of "kinds"? Actually, yes. Research from the field of systematics and phylogenetics could, and does disprove the existence of "kinds". A creationist ought to admit the validity of phylogenetic analyses that identify monophyletic clades (roughly the evolutionary equivalent to a monobaramin) of closely related species within a holobaramin (roughly equivalent to a genus) since under both the creation model and the theory of evolution all of these species have shared a recent common ancestor. Differences in DNA sequences between the species that have accumulated since they developed reproductive isolation are the information that molecular phylogenetic analyses use to establish historical patterns of evolution. The more time that has passed since two species have shared a recent common ancestor, the more dissimilar their DNA should be. Under the creation model, closely related species descended from the same ancestral "kind" should have DNA sequences that show the patterns of speciation since diverging from the parent species. Phylogenetic analyses of these DNA sequences should show which species are more related to which species within the "kind", but this should only hold true when comparing DNA from species within a single holobaramin, or "kind". Such patterns should not and need not exist when comparing species from different holobaramins. In other words, phylogenetic analyses should nicely elucidate the relationships within a single holobaramin. However, when you compare DNA sequences from species in different holobaramins, there should be no reason that a pattern indicating further descent from a more distant common ancestor should exist. In fact, under the creation model, a phylogenetic analysis of sequences from a canid (dog) and felid (cat) holobaramin should not indicate they shared a recent common ancestor with each other any more recently than either did with a sponge, fern, Paramecium or primate. Quite simply, the analysis should spit out a nonsense phylogeny for organisms compared above the level of the "kind", and it should not be consistent with relationships proposed by evolutionary biologists based on the premise that all life descended from a common ancestor and was determined using independent evidences from the fossil record, comparative morphology, physiology, behavior, and biogeography. Unfortunately for creationists, this is not the case.
I will illustrate this using two case studies. The first is phylogenetic analyses of "cats" (Family: Felidae). Friar states that:
"Cats" constitute another holobaramin with monobaraminic branches for the lion and the tiger, for the pumas, another for the lynx, domestic cats, etc.
Friar does not provide any reference(s) for these claims, so it is either based on common sense or from the results of an uncited study conducted by another creationist. In any event, Friar is claiming that felids form a single holobaramin (i.e. they are all descended from a single common ancestor – the felid "kind"). Further, within this holobaramin, there are monobaramins (monophylies) containing the lion and tiger, another for the pumas, another for the lynx and another for domestic cats. Consider a phylogeny of felids based on the combined data from 3 DNA regions (Figure 1). We can see that the results of this analysis support the general descriptions offered by Friar, and that all felids have descended from a common felid ancestor. So far so good for the creationists – and we have demonstrated that phylogenetic analyses work within the level of a "kind".
The question now becomes – what if we compare DNA sequences from felids along with other mammals? Under the creation model, there is no reason to expect that, for example, otters should share a more recent common ancestor with weasels than either does with any other group – or whether dogs should share a more recent common ancestor with seals than with cats. Under the creation model, phylogenetic methods should not reveal any meaningful patterns when considering sequence data from different holobaramins with the Order Carnivora. And yet they do – patterns that are consistent with proposed relationships based on morphology and other molecular studies (Figure 2). And if you search even more broadly considering placental mammals, you find the same branching patterns you found below the level of "kinds" indicating common ancestry, and consistent with relationships proposed based on other data (Figure 3).
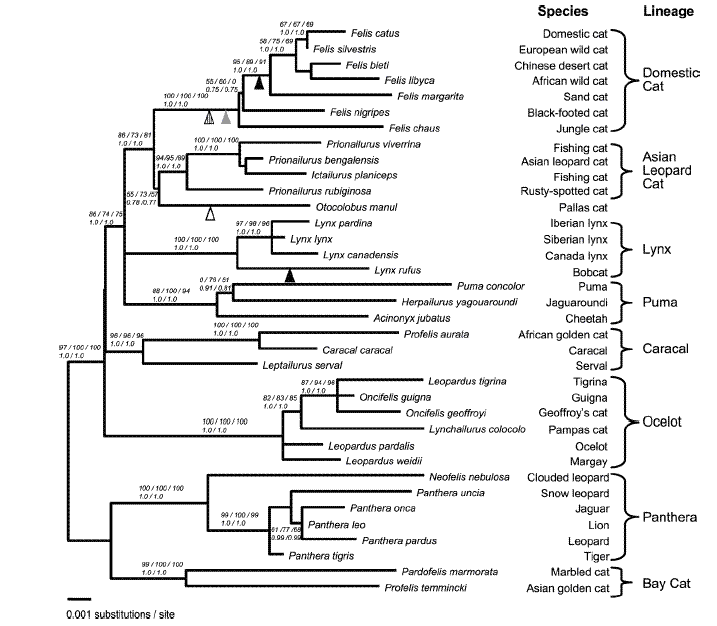
Figure 1. Phylogenetic tree of combined data from three Y-chromosome genes SMCY, UBE1Y, and ZFY (2462 bp) sequenced from 36 species of felids. Source: Pecon-Slattery et al.)
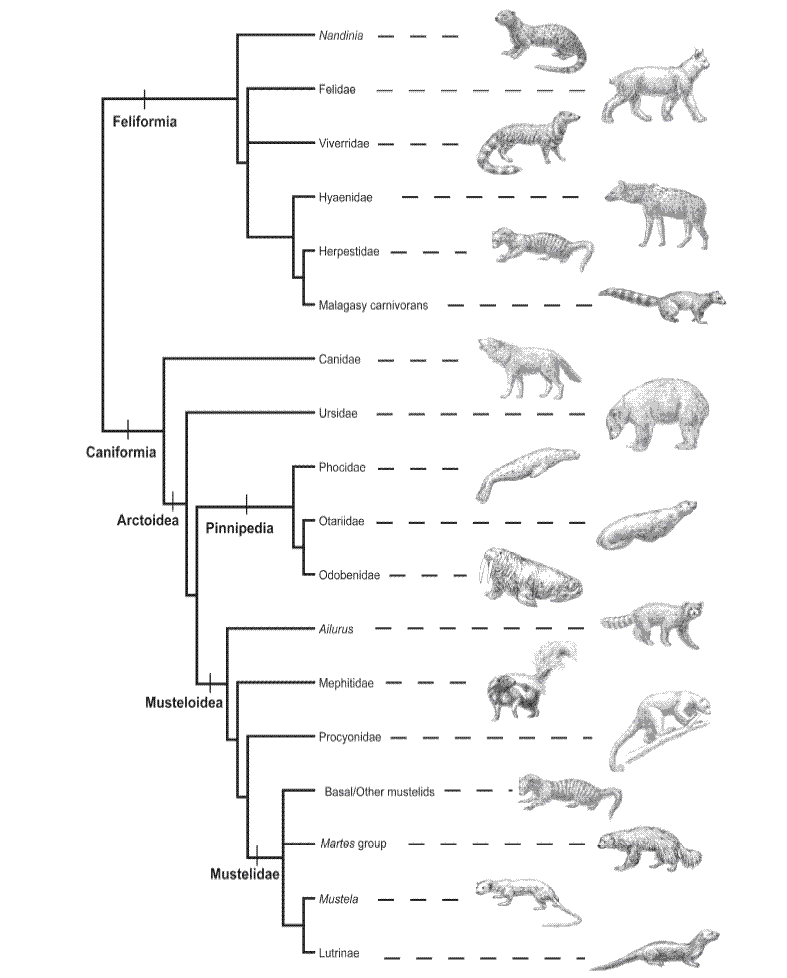
Figure 2. A schematic cladogram representing the major evolutionary relationships recovered in an analysis of Carnivora. (Source: Flynn et al. 2005).
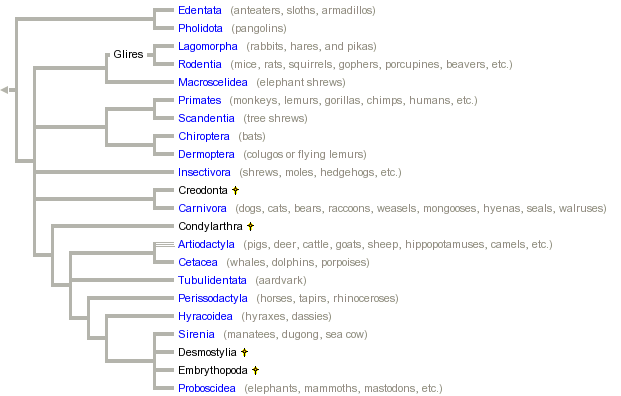
Figure 3. Phylogeny of placental mammals. (Source: http://www.tolweb.org/Eutheria/15997)
Another study on the bariminology of snakes (Hennigan, 2005) attempts to identify "kinds" of snakes using hybridization data. Hennigan chooses to use the presence of documented examples of hybrids forming between snake species to determine the monobaramins within the snake "kind". Hennigan's first step was "to determine true discontinuity". Friar (2000) states that:
it is important to emphasize that the strictly empirical component of baraminology is discontinuity systematics which can be utilized by itself without any reference to religious literature.
Yet Hennigan's first step involves the use of the so-called Discontinuity Matrix to establish discontinuity – and the first two questions in this matrix are:
does the Bible claim discontinuity for snakes?
and
does the bible imply discontinuity for snakes?
Clearly this is directly violating the claim that discontinuity systematics can be practiced without any reference to religious literature. After (surprisingly) establishing discontinuity, Hennigan proceeds with his analysis of hybridization records. Hennigan offers no explanation for why he chose hybridization over all other sources of data in his analysis besides this statement (which had no reference to any literature to support it):
Hybridization suggests a close biochemical relationship between two organisms and is an important additive evidence for ancestry within a "kind" in the identification of monobaramins.
He admits that:
just because two organisms are unable to hybridize does not mean they are unrelated.
and so he weakly offers a caveat that other sources of data will be considered and he plans to treat the organisms "holistically". Once again, the creationists fail to invoke any set of objective, rigorous and quantitative methods in their analysis.
Hennigan identified roughly six monobaramins, but his results are not strongly consistent with the results of phylogenetic analyses using molecular data (Figure 4). Nevertheless, the results of the phylogenetic analyses from Heise et al. (1995) do effectively describe relationships of snakes, and indicate that they are monophyletic – and they could offer an answer to Hennigan's question of whether snakes are descended from one, or several ancestral "kinds". The question is whether relationships continue to be described when species other than snakes are considered. Hennigan hypothesizes:
that snakes are discontinuous with any other group and are therefore an apobarimin.
However, when DNA sequences from snakes are analyzed together with other squamates (reptiles) the results not only yield a monophyletic clade for snakes, but a monophyletic clade consisting of all squamates including snakes thereby demonstrating that these different "kinds" have shared a common ancestor (Figure 6).
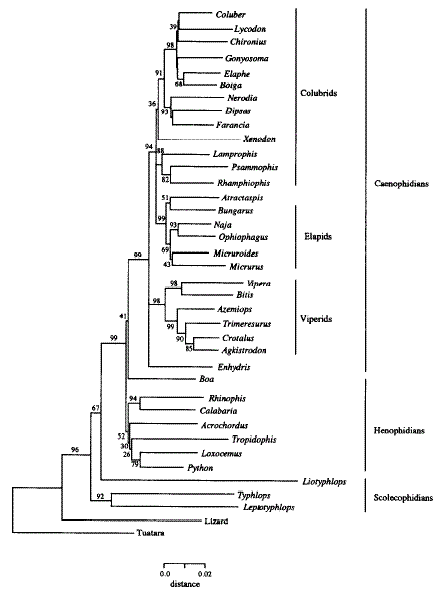
Figure 4. Phylogenetic tree of snakes based on mitochondrial 12s and 16s rRNA sequence data (8 18 aligned sites). (Source: Heise et al.1995).
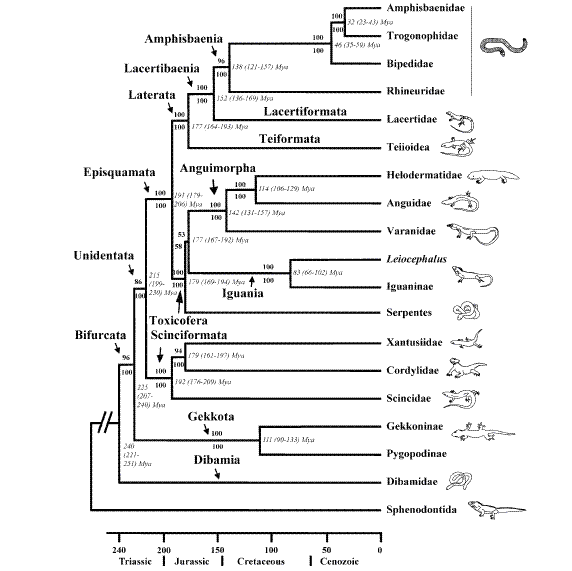
Figure 5. Phylogenetic relationships of squamates inferred from DNA sequences of nine nuclear protein-coding genes (C-mos, RAG1, RAG2, R35, HOXA13, JUN, α-enolase, amelogenin, and MAFB, totaling 6192 bp).(Source: Vidal & Hedges 2005).
In conclusion, phylogenetic methods which have often detected patterns of relationships predicted by creationists below the level of a "kind" which indicate species have descended from a common ancestor, continue to detect patterns above the level of a "kind". The patterns are logical, consistent with relationships proposed by biologists based on other data, and indicate that life has descended from common ancestors beyond the level of the "kind". Hennigan (2005) states:
Creation theory… postulates a "forest" of organisms in which each "tree" began with an originally created pair designed with vast genetic potential for variation but discontinuous with (not related to) the other created "kinds".
The pattern should look similar to the trees in Figure 6. Below the creation event in Figure 6, no phylogenetic patterns should exist – no logical relationships should be found among the different kinds; yet this is precisely what we find. In other words, there is no demarcation at the level of holobaramins, and "kinds" do not exist.
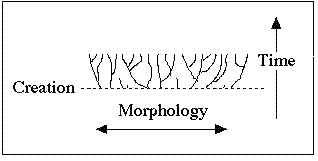
Figure 6. The True Creationist 'Orchard' (Source: http://www.nmsr.org/sarfati.htm)
Sloppy writing and structure
The manuscript itself is very poorly written and the structure and composition leave much to be desired. There is a reference in the text not found in the literature cited section (Gaffney and Meylan (no date provided)). Unconventionally, Friar cites personal communications in the literature cited section (e.g. Cavanaugh, 1999–2000), figures are not numbered or titled, sentence structure is often poor (e.g. "Scripture claims (used in baraminology but not in discontinuity systematics)."), there are occasional misspellings or improper use of capital or lower case letters (ReMine vs. Remine, caucasian), and punctuation errors ("For example the current Order Primates…"). The unlabeled figures do not even appear to be the figures that are referenced in the text. This is important for two reasons; first, creationists often charge the scientific community with bias and an unwillingness to consider their manuscripts for publication. Aside from the fact that their research is unscientific, and as such has no place in the scientific literature, if this paper is indicative of the quality of manuscript that would be submitted for peer review, I can say certainly that on the basis of the style and writing alone it would be rejected by the editor without being sent for peer review. What is more shocking is that this is not a draft copy – this article was actually published in their own journal. So the second reason the poor quality of the writing is significant is that creationists often cite their own work (as Friar has done abundantly in this paper) and insist that it be taken as seriously as research published in mainstream science journals. That a paper like this could make it through peer review (assuming they use peer review) is astonishing.
Human "Kind"
Paleontological and molecular evidences have demonstrated that humans have shared a common ancestor with other primates (Jaeger & Marivaux 2005, Poux & Douzery 2004), yet this will never be reconciled with creationist baraminology given the primacy of man in God's creation. Instead, Friar states clearly and unequivocally that humans form a holobaramin with the various races at the branches. He even goes so far as to make the following statement:
A member of any of these races potentially would be inter-fertile with a spouse of the opposite sex from any other race.
Potentially??? Friar seems to want to apply Hennigan's hybridization rules used on different species of snakes to humans in order to establish the existence of human races. Friar is not just stating the obvious – that members of different "races" marry and bear healthy children is an absurdly obvious observation – but it ignores a large body of research on human genetic variation. For example, Keita et al. (2004) state:
The within- to between-group variation is very high for genetic polymorphisms (~85%). This means that individuals from one "race" may be overall more similar to individuals in one of the other "races" than to other individuals in the same "race".
In short, regardless of whether races exist at the genetic level in humans or not, humans, alone, do not form a holobaramin when the paleontological and molecular evidences for primates are considered more broadly. But because of the strict limits imposed on baraminology, humans can never be included in another group with other primates because it contradicts scripture.
Specific Criticisms
The stage now was set, and subsequently billions of science students would play their part upon the stage of life and during that time sense the excitement of discovering God's message in nature (Psalm 19:1; Romans 1:19,20; and see ReMine, 1993).
Besides being a little melodramatic, it seems a little odd to me to support a statement in a "scientific" journal article simultaneously with scriptural references and references to other articles.
For example there is phyletic or evolutionary systematics (or Darwinian systematics) in which macroevolution (involving large changes) is assumed.
Actually, the word phyletic is usually applied to the mode or tempo of evolution (viz. phyletic gradualism). As such, it is more appropriately associated with microevolutionary changes that, over long periods of time, result in macroevolutionary changes. A phyletic method of classification attempts to group organisms based on a shared common ancestor. The word is not generally synonymous with evolutionary systematics, except in the sense that systematics is the study of evolutionary relationships among organisms and the characters that define them, and it assumes all life has descended from common ancestry. Phrases like "evolutionary systematics" and "Darwinian systematics" are rarely, if ever used. I believe that Friar is intending to describe phylogenetics, of which cladistics is the predominant method invoked. Friar, himself, seems a little confused about phylogenetics adding that:
… there is the cladistic approach which focuses on common or so-called "shared/derived" characters.
While he is right that cladistics uses shared, derived characters in their methods, cladistics is a method of phylogenetic analysis – not a separate discipline as Friar implies.
Most importantly, popular methodology in phyletic, phenetic and cladistic procedures all have been adapted to macroevolutionary theorizing.
Friar does not elaborate on the significance of this statement.
The earliest methods were phenetic which conferred the distinct advantage of being objectively observable and most verifiable.
I am not certain what this statement means. If he means that phenetic analyses are superior because they use "observable" phenotypic characters, well so do cladistic analyses. It is further unclear how they are verifiable. Verifiable in relation to what? Phenetic methods are based on overall similarity, and can be used to classify virtually anything – you could use it to group cars (V6 or V8? Two-door or four door?) or shoes in your closet (laces or no-laces?). But just as two shoes that have many traits in common are not descended from a common ancestor, neither may two demonstrably similar organisms. It is likely that two organisms that are phenotypically similar share a recent common ancestor, but not necessarily since the similar structures may be the products of convergent evolution. Phenetic methods cannot discern this, and as such, I am not certain what Friar means by "verifiable".
Scientists who have preferred something other than a macroevolutionary framework recently have developed what they believe to be a more realistic systematics based upon the discontinuities or typology found in nature.
No reference is provided for this vital assumption.
But the baraminologist maintains that thinking about phylogeny should not extend beyond convincing evidence, and that scientists should be relieved of their sense of obligation philosophically to construct extensive phylogenies (evolutionary trees) in the absence of compelling facts.
One is left to wonder to what "convincing evidence" Friar is referring. He seems to be broadly dismissing scientific research in the fields of paleontology, embryology, comparative morphology and physiology, genetics and molecular biology as insufficient evidences that life shares common ancestry beyond the level of "kinds". To ignore the evidences in these fields – worse, to consider them not compelling is remarkably glib.
In addition, systematic studies on particular monobaramins where there has been diversification (as the appearance of more recognizable species, subspecies, varieties, etc.) the research on a monobaramin would not differ essentially depending on the systematic philosophy of the investigators.
Someone please have that sentence taken out behind the woodshed and thoroughly flogged. Still, he seems to be admitting that phylogenetic methodologies within the level of a "kind" could be applied equally as baraminology. That these methods continue to work above the level of the "kind" seems not to impress him, but implies that they should not be taken seriously above the level of "kinds" due to the misguided philosophy of the evolutionary biologist.
The group of all humans and all turtles also would be apobaraminic because no human or turtle shares ancestry with any non-human or non-turtle organisms.
This is a clearly false statement, and Friar has chosen to ignore rather than refute the evidence from the scientific literature on the phylogenetics and systematics of turtles, squamates and primates.
So a group containing the human holobaramin and the chimp-gorilla holobaramin would be apobaraminic (see Figure 7A). Further, a collection of the human holobaramin with any or all the other primate holobaramins would be apobaraminic. No member of any of these holobaramins would share any ancestry with a member of any of the other holobaramins within or even outside this apobaramin.
Yet again, Friar makes an unfounded remark and blithely ignores the scientific literature on primate evolution.
For people reared on an evolutionary diet the above menu can be difficult to swallow and digest because students of biology have been taught to think genetic relationship rather than genetic discontinuity.
Um. Because there is evidence for genetic relationships, and NO evidence for genetic discontinuity.
We are not "cutting and pruning" the data. Rather, we leave the data precisely where it is.
The word "data" is plural. "Datum" is singular. Friar, a biologist, should know this. Thus, the sentence should read "Rather, we leave the data precisely where they are". Regardless, it is not clear what he is saying in that sentence.
In other words the scientist is iterating tentative taxonomies by increasing or decreasing sizes of the branches to arrive at the best approximation of reality.
In other words, the creationist is making it up as they go along.
For those who have been steeped in Linnaean taxonomy and evolutionary thinking, discontinuity systematics may appear to be a preposterous proposal.
That's the understatement of the year.
Literature Cited
Excoffier, L. G. Laval, and S. Schneider (2005) Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online 1:47-50
Flynn, J.J., Finarelli, J.A., Zehr, S., Hsu, J., and Nedbal, M.A. (2005). Molecular Phylogeny of the Carnivora (Mammalia): Assessing the Impact of Increased Sampling on Resolving Enigmatic Relationships. Syst. Biol. 54(2):317–337
Philip J. Heise, P.J., Maxson, L.R., Dowling, H.G. and Hedges, S.B. (1995). Higher-Level Snake Phylogeny Inferred from Mitochondrial DNA Sequences of 12s rRNA and 16s rRNA Genes. Mol. Bid. Ed. 12(2):259-265.
Hennigan, T. (2005). An Initial Investigation into the Baraminology of Snakes: Order—Squamata, Suborder Serpentes. CRSQ 42:153-160
Hillis, D.M., C. Moritz and B. Mable (eds). 1996. Molecular Systematics 2nd. ed., Sinauer Assoc., Sunderland, MA.
Penny, D., Hendy, M.D. and Steel, M.A. (1992). Progress with methods for constructing evolutionary trees. Trends in Ecology and Evolution 7: 73-79
Poux, C. & Douzery, E.J.P. (2004). Primate Phylogeny, Evolutionary Rate Variations, and Divergence Times: A Contribution From the Nuclear Gene IRBP. American Journal of Physical Anthropology 124:1–16.
Rambaut A & Grassly NC (1997). Seq-Gen: an application for the Monte Carlo simulation of DNA sequence evolution along phylogenetic trees Comput Appl Biosci 13, 235-238
Saux, C., Fisher, B.L., and Spicer, G.S. (2004). Dracula ant phylogeny as inferred by nuclear 28S rDNA sequences and implications for ant systematics (Hymenoptera: Formicidae: Amblyoponinae) Molecular Phylogenetics and Evolution 33:457–468
Vidal, N. and Hedges, S.B. (2005). The phylogeny of squamate reptiles (lizards, snakes, and amphisbaenians) inferred from nine nuclear protein-coding genes. C. R. Biologies 328: 1000–1008.
Weiss, G. and von Haesler, A. (2003). Testing substitution models within a phylogenetic tree. Mol. Biol. Evol. 20(4):572-578
![]()